
Contents
Lorsque le journaliste Roger Lewin a baptisé en 1987 l’ancêtre maternel commun à tous les peuples vivant aujourd’hui « Eve Mitochondriale », il a évoqué l’histoire de la création, celle d’une femme qui était notre mère à tous et dont les descendants se sont dispersés sur la terre. Le nom a capté l’imagination collective et est toujours utilisé non seulement par le public mais aussi par de nombreux scientifiques pour désigner cet ancêtre maternel commun.
Mais le nom a été plus trompeur qu’utile. Il a favorisé l’impression erronée que tout notre ADN provient de deux ancêtres précisément et que pour connaître notre histoire, il suffirait de suivre simplement la lignée purement maternelle représentée par l’ADN mitochondrial, et la lignée purement paternelle représentée par le chromosome Y.
Inspiré par cette possibilité, le « Projet Genographic » de la National Geographic Society, qui a débuté en 2005, a recueilli des données sur l’ADN mitochondrial et le chromosome Y auprès de près d’un million de personnes de divers groupes ethniques . Mais le projet était déjà dépassé avant même d’avoir commencé. Il a été largement récréatif et n’a produit que peu de résultats scientifiques intéressants. Dès le début, il était clair que la plupart des informations sur le passé humain présentes dans l’ADN mitochondrial et les données du chromosome Y avaient déjà été exploitées, et que des histoires bien plus riches étaient enfouies dans l’ensemble du génome. La vérité est que le génome contient les histoires de nombreux ancêtres différents – des dizaines de milliers de lignées généalogiques indépendantes, et pas seulement les deux dont l’histoire peut être retracée grâce au chromosome Y et à l’ADN mitochondrial.
Pour comprendre cela, il faut réaliser qu’au-delà de l’ADN mitochondrial, le génome n’est pas une séquence continue provenant d’un seul ancêtre, mais plutôt une mosaïque. Quarante-six des tuiles de la mosaïque sont en quelque sorte des segments d’ADN chromosomiques qui sont physiquement séparés dans la cellule. Un génome est constitué de vingt-trois chromosomes, et comme une personne porte deux génomes, un de chaque parent, le nombre total est de quarante-six.
Chromosomes. ( peterschreiber.media / Adobe)
Mais les chromosomes eux-mêmes sont des mosaïques de tuiles encore plus petites. Par exemple, le premier tiers d’un chromosome qu’une femme transmet à son ovule peut provenir de son père et les deux derniers tiers de sa mère, résultat de l’épissage des copies de ce chromosome de son père et de sa mère dans ses ovaires. Les femmes créent en moyenne environ quarante-cinq nouvelles épissures lorsqu’elles produisent des ovules, tandis que les hommes en créent environ vingt-six lorsqu’ils produisent des spermatozoïdes, soit un total d’environ soixante et onze nouvelles épissures par génération. Ainsi, en remontant chaque génération plus loin dans le passé, le génome d’une personne est dérivé d’un nombre toujours croissant de fragments ancestraux épissés ensemble.
Cela signifie que nos génomes renferment une multitude d’ancêtres. Le génome de toute personne est dérivé de 47 segments d’ADN correspondant aux chromosomes transmis par la mère et le père, plus l’ADN mitochondrial. Il y a une génération, le génome d’une personne est dérivé d’environ 118 (47 plus 71) segments d’ADN transmis par ses parents. Il y a deux générations, le nombre de segments d’ADN ancestraux s’élève à environ 189 (47 plus 71 plus 71 autres) transmis par quatre grands-parents. Si l’on remonte encore plus loin dans le temps, on constate que l’augmentation supplémentaire des séquences d’ADN ancestrales à chaque génération est rapidement dépassée par le doublement du nombre d’ancêtres. Dix générations plus tôt, par exemple, le nombre de segments d’ADN ancestraux est d’environ 757, mais le nombre d’ancêtres est de 1 024, ce qui garantit que chaque personne a plusieurs centaines d’ancêtres dont elle n’a reçu aucun ADN. Sur vingt générations, le nombre d’ancêtres est presque mille fois supérieur au nombre de segments d’ADN ancestraux dans le génome d’une personne. Il est donc certain que chaque personne n’a pas hérité d’ADN de la grande majorité de ses ancêtres actuels.

Assemblage de brins d’ADN à partir de différents éléments. ( Tatiana Shepeleva / Adobe )
Ces calculs signifient que la généalogie d’une personne, telle que reconstruite à partir de documents historiques, n’est pas la même que son héritage génétique. La Bible et les chroniques des familles royales mentionnent qui a engendré qui sur des dizaines de générations. Pourtant, même si les généalogies sont exactes, la reine Elizabeth II d’Angleterre n’a presque certainement pas hérité d’ADN de Guillaume de Normandie, qui a conquis l’Angleterre en 1066 et qui serait son ancêtre vingt-quatre générations plus tôt. Cela ne signifie pas que la reine Elizabeth II n’a pas hérité de l’ADN d’ancêtres aussi anciens, mais on s’attend à ce que seuls 1 751 de ses 16 777 216 ancêtres généalogiques du vingt-quatrième degré lui aient apporté de l’ADN. C’est une si petite fraction que la seule façon pour William d’être plausible son ancêtre génétique est qu’il soit son ancêtre généalogique dans des milliers de lignées différentes, ce qui semble peu probable même en considérant le haut niveau de consanguinité dans la famille royale britannique.
En remontant plus loin dans le temps, le génome d’une personne se retrouve dispersé dans des étendues d’ADN de plus en plus ancestrales, réparties sur un nombre toujours plus grand d’ancêtres. En remontant cinquante mille ans dans le passé, notre génome est dispersé en plus de cent mille portions d’ADN ancestrales, plus que le nombre de personnes qui vivaient dans une population à cette époque. Nous héritons donc de l’ADN de presque tous les membres de notre population ancestrale qui ont eu un nombre important de descendants à des époques aussi lointaines.
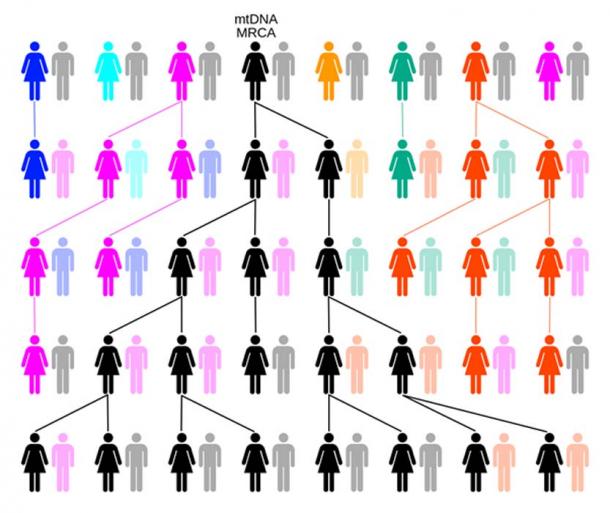
Par dérive ou sélection aléatoire, la lignée féminine remontera jusqu’à une seule femelle, comme l’Eve mitochondriale. Dans cet exemple, sur cinq générations, les couleurs représentent les lignées matrilinéaires éteintes et le noir la lignée matrilinéaire descendant de l’ADN mitochondrial MRCA. (ChrisTi / CC BY-SA 3.0 )
Il y a cependant une limite aux informations que la comparaison des séquences du génome fournit sur le temps profond. À chaque endroit du génome, si nous remontons suffisamment loin dans le passé, nous atteignons un point où tout le monde descend du même ancêtre, au-delà duquel il devient impossible d’obtenir des informations sur le temps profond à partir de la comparaison des séquences d’ADN des personnes vivant aujourd’hui.
De ce point de vue, l’ancêtre commun à chaque point du génome est comme un trou noir en astrophysique, auquel aucune information sur le temps profond ne peut échapper. Pour l’ADN mitochondrial, ce trou noir se produit il y a environ 160 000 ans, la date de « l’Eve mitochondriale ». Pour la grande majorité du reste du génome, le trou noir se produit entre cinq millions et un million d’années, et donc le reste du génome peut fournir des informations sur un temps bien plus profond que ce qui est accessible par l’analyse de l’ADN mitochondrial. Au-delà de cela, tout s’assombrit.
Le pouvoir de tracer cette multitude de lignées pour révéler le passé est extraordinaire. Dans mon esprit, lorsque je pense à un génome, je ne le vois pas comme une chose du présent, mais comme une tapisserie de fils profondément enracinés dans le temps, une tapisserie constituée de lignes de descendance et de séquences d’ADN copiées de parent à enfant remontant dans le passé lointain. En remontant, les fils s’enroulent autour d’ancêtres toujours plus nombreux, fournissant des informations sur la taille et la sous-structure de la population à chaque génération.
Quand on dit qu’un Afro-Américain a 80 % d’ascendance ouest-africaine et 20 % d’ascendance européenne, par exemple, on affirme qu’il y a environ cinq cents ans, avant les migrations et les mélanges de populations précipités par le colonialisme européen, 80 % des fils ancestraux de la personne résidaient probablement en Afrique de l’Ouest et le reste en Europe. Mais de telles affirmations sont comme les images fixes d’un film, capturant un point dans le passé. Une perspective tout aussi valable est qu’il y a cent mille ans, la grande majorité des lignées d’ancêtres afro-américains, comme celles de tout le monde aujourd’hui, se trouvaient en Afrique.

Visualisation des données génomiques. Séquence du génome de l’ADN, carte génétique médicale. ( MicroOne / Adobe )
L’histoire racontée par les multitudes dans nos génomes
En 2001, le génome humain a été séquencé pour la première fois – ce qui signifie que la grande majorité de ses lettres chimiques ont été lues. Environ 70 % de la séquence provient d’un seul individu, un Afro-Américain, mais certaines lettres proviennent d’autres personnes. En 2006, des entreprises ont commencé à vendre des robots qui ont réduit le coût de la lecture des lettres de l’ADN de plus de dix mille fois et bientôt de cent mille fois, rendant ainsi économique la cartographie du génome de beaucoup plus de personnes. Il est ainsi devenu possible de comparer les séquences non seulement de quelques endroits isolés, comme l’ADN mitochondrial, mais de tout le génome. Cela a permis de reconstruire les dizaines de milliers de lignées ancestrales de chaque personne. Cela a révolutionné l’étude du passé. Les scientifiques ont pu rassembler des ordres de grandeur supplémentaires de données et vérifier si l’histoire de notre espèce suggérée par l’ensemble du génome était la même que celle racontée par l’ADN mitochondrial et le chromosome Y.
En 2011, un article de Heng Li et Richard Durbin a montré que l’idée que le génome d’une seule personne contienne des informations sur une multitude d’ancêtres n’était pas seulement une possibilité théorique, mais une réalité. Pour déchiffrer l’histoire profonde d’une population à partir de l’ADN d’une seule personne, Li et Durbin ont exploité le fait que toute personne porte en fait non pas un mais deux génomes : un de son père et un de sa mère. Ainsi, il est possible de compter le nombre de mutations séparant le génome qu’une personne reçoit de sa mère et le génome qu’elle reçoit de son père afin de déterminer quand ils ont partagé un ancêtre commun à chaque endroit.
En examinant l’éventail des dates où ces ancêtres ont vécu – en traçant les âges de cent mille Adams et Eves-Li et Durbin – on a pu établir la taille de la population ancestrale à différentes époques. Dans une petite population, il y a de fortes chances que deux séquences du génome choisies au hasard dérivent de la même séquence du génome parent, car les individus qui les portent partagent un parent. Toutefois, dans une population importante, cette probabilité est bien plus faible. Ainsi, les périodes du passé où la taille de la population était faible peuvent être identifiées sur la base des périodes du passé où une fraction disproportionnée des lignées ont la preuve de partager des ancêtres communs.
Walt Whitman, dans le poème « Song of Myself », a écrit : « Est-ce que je me contredis ? / Très bien, alors je me contredis, / (je suis grand, je contient des multitudes). » Whitman aurait tout aussi bien pu parler de l’expérience Li et Durbin et de sa démonstration que toute l’histoire d’une population est contenue dans une seule personne, comme le révèle la multitude d’ancêtres dont les histoires sont enregistrées dans le génome de cette personne.
L’étude de Li et Durbin a révélé de façon inattendue qu’après la séparation des populations non africaines et africaines, il y a eu une longue période dans l’histoire commune des non-Africains où les populations étaient peu nombreuses, comme le montrent les preuves de nombreux ancêtres communs étalés sur des dizaines de milliers d’années. Un « goulot d’étranglement » commun aux non-Africains – lorsqu’un petit nombre d’ancêtres a donné naissance à un grand nombre de descendants aujourd’hui – n’est pas une découverte nouvelle. Mais avant les travaux de Li et Durbin, on ne disposait pas de bonnes informations sur la durée de cet événement, et il semblait plausible qu’il ait pu se produire sur quelques générations seulement – par exemple, un petit groupe de personnes traversant le Sahara pour se rendre en Afrique du Nord, ou de l’Afrique vers l’Asie .
Les preuves de Li et Durbin d’une période prolongée de faible population étaient également difficiles à concilier avec l’idée d’une expansion imparable de l’homme moderne à la fois à l’intérieur et à l’extérieur de l’Afrique il y a environ cinquante mille ans. Notre histoire n’est peut-être pas aussi simple que l’histoire d’un groupe dominant qui a connu un succès immédiat partout où il est allé.
Image du haut : Adam et Eve (rudall30 / Adobe Stock)
Par David Reich
© [Oxford University Press]. Extrait de Qui nous sommes et comment nous sommes arrivés ici : Ancient DNA and the new science of the human past par David Reich, publié par Oxford University Press, disponible en format relié, livre de poche et livre électronique, 10,99
.








